Interaction chêne/herbacées : la faim vient en mangeant
Publié par Journal Décoder, le 21 juillet 2023 2.3k

Antoine Vernay1 (enseignant-chercheur) et la classe de 2nd 2 du lycée Pierre Brossolette de Mme Christine Louis2 et Mr Benoit Le Hir2 (la liste des élèves est mentionnée en fin d’article)
Article original/Original article: Vernay, Antoine, Philippe Malagoli, Marine Fernandez, Thomas Perot, Thierry Améglio, et Philippe Balandier. « Improved Deschampsia cespitosa growth by nitrogen fertilization jeopardizes Quercus petraea regeneration through intensification of competition ». Basic and Applied Ecology 31 (1 septembre 2018): 21‑32.
Institution: 1Univ Lyon, Université Claude Bernard Lyon 1, CNRS, ENTPE, UMR 5023 LEHNA, F-69622, Villeurbanne,France
2Lycée Pierre Brossolette, 1 Esplanade Agnès Varda, 69100 Villeurbanne
Résumé :
La régénération des chênaies est compliquée du fait de la présence d’herbacées comme la canche.
La disponibilité de plusieurs ressources est en jeux pour expliquer les échecs de régénération avec les herbacées mais peu d’étude ont pu quantifier et caractériser ces interactions en faisant varier plusieurs niveaux de ressource.
Lors de plusieurs expérimentations, les scientifiques ont fait varier plusieurs facteurs comme la lumière et l’azote. Ils ont donc planté de jeunes plants de chêne et de canche en les mettant en interaction ou non. Ces expériences ont permis ainsi de comprendre comment réagissaient les végétaux à différents niveaux de disponibilité de ressources et à la compétition.
L’étude montre que, selon le niveau de ressource de lumière et d’azote, les interactions entre les deux plantes varient. Dès qu’il y a une ressource abondante, la plante est beaucoup plus en compétition que le chêne. C’est la double peine pour le chêne, celui-ci pourrait faciliter le développement de la canche en lui fournissant une source d’azote (rhizodépôt) et dans le même temps la canche réduit la croissance du chêne en captant la plupart de l’azote.
Mots clés : canche ; chêne ; compétition pour les ressources ; gestion forestière ; stratégie écologique
I. Introduction
Plusieurs théories en écologie tentent d’expliquer les échanges entre les plantes pour les ressources. Certaines accaparent rapidement beaucoup de ressources et se développent rapidement, on parle de stratégie acquisitrice. D’autres ont tendance à recycler et stocker les ressources induisant une croissance plus lente, on parle de stratégie conservatrice. Ces comportements des plantes peuvent varier selon les conditions environnementales, c’est ce qu’on appelle la plasticité phénotypique (capacité d’un organisme à exprimer différents caractères pour un même génotype). Dans un même milieu, des plantes voisines consomment les mêmes ressources et peuvent donc entrer en compétition pour celles-ci, lorsque la présence d’une plante A induit une baisse de performance d’une plante B (en réduisant considérablement la disponibilité d’une ressource par exemple, diminuant la croissance de la plante B) mais la plante A peut aussi faciliter la croissance de la plante en B en créant de l’ombrage par exemple et diminuant l’évaporation de l’eau ce qui diminue le stress hydrique (manque d’eau pour la plante, qui se traduit par une plus faible croissance).
La capacité d’une plante à survivre dans un environnement dépend donc de la stratégie écologique des plantes voisines et des conditions environnementales qui impactent le comportement des plantes. On peut alors différencier l’intensité, l’effet de la présence d’une plante sans tenir compte de l’environnement et l’importance des relations par rapport aux stress dues aux conditions environnementales (Brooker et al., 2005).
Tenir compte des stress multiples du milieu, des stratégies des plantes voisines, et de la capacité de la plante à modifier son comportement, rend plus compliqué mais aussi plus proche de la réalité notre compréhension du fonctionnement des plantes dans leur milieu que de ne considérer qu’un facteur pour expliquer les interactions. Cette difficulté augmente aussi en considérant la réponse des plantes selon plusieurs facteurs qui changent : il est plus facile de ne faire varier qu’un seul facteur, par exemple la lumière, pour expliquer le comportement des plantes mais dans la réalité, ce sont plusieurs facteurs qui varient de manière simultanée (lumière, eau, nutriments, température, …).
Dans cette étude, les chercheur·se·s ont choisi de faire varier deux facteurs, la lumière (L) et l’azote du sol (N) afin de comprendre leurs impacts sur les interactions entre des jeunes plants de chênes sessiles (Quercus petraea (Matt.) Liebl., 1784) et de la canche cespiteuse (Deschampsia cespitosa (L.) P.Beauv., 1812). Ces deux espèces sont parmi les plus communes des chênaies tempérées françaises, le chêne sessile ayant une bonne résistance à la sécheresse, il devrait voir son aire de répartition augmenter du fait du dérèglement climatique (IGN, 2016). La canche est une Poacée qui induit beaucoup de mortalité chez les jeunes chênes du fait de leur stratégie très acquisitrice. Les hypothèses des chercheur·se·s prévoient une atténuation de la compétition entre ces deux espèces dans les cas de faible disponibilité en lumière, du fait d’une moins bonne performance de croissance des canches. Nous pensons également que le chêne allouera plus de ressources aux racines dans les environnements avec peu de N et plus de ressources aux feuilles dans les cas de faible lumière (augmentation des performances des organes liés à l’acquisition des ressources limitantes), ce qui lui permettra de faire face à la stratégie d’appropriation de la canche à court terme.
II. Matériels et méthodes
L’expérience a eu lieu dans une serre à Clermont-Ferrand entre décembre 2014 et juin 2015. 120 chênes d’un an ont été plantés dans des pots de 20L. Des plants de canche (herbacé), découpés en thalles* (lexique en fin d’article) ont été récoltés dans une forêt de chêne de l’Allier puis replantés par nombre de trois (correspondant à la densité retrouvée en chênaie) dans 80 pots contenant des chênes. 40 pots ne contenaient alors qu’un chêne et 40 autres pots uniquement trois thalles de canche. Pour chacun des traitements de diversité, la moitié des pots a été mise sous ombrière, l’autre est restée sous la lumière de la serre. Pour les pots bispécifiques (chêne +canche, traitement « diversité »), 40 seront sous ombrière et 40 en pleine lumière. Dans chacune des conditions « diversité × lumière », la moitié des pots ont reçu une fertilisation (action de fertiliser le sol) en N en 3 fois, équivalente à la fin à 89kg.ha-1. Les autres n’ont pas reçu de fertilisant (produits destinés à assurer ou à améliorer la nutrition des végétaux et les propriétés des sols.) (Figure 1).
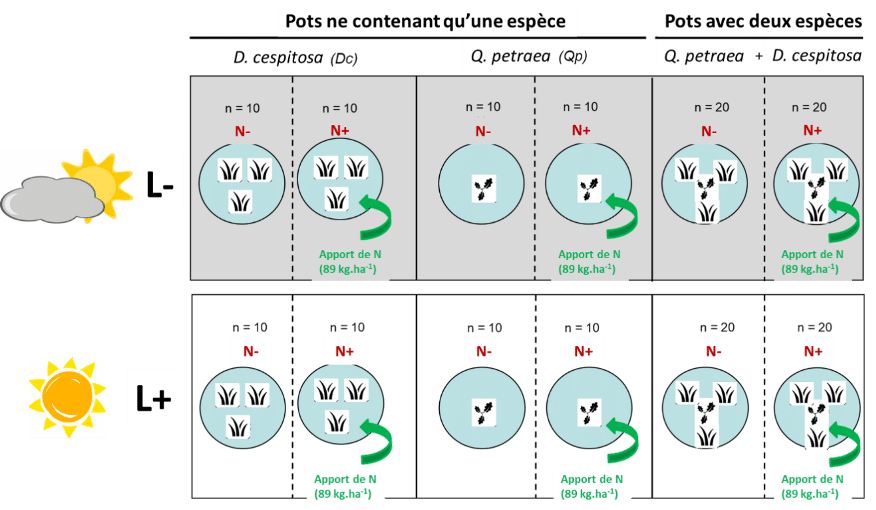
Figure 1 : Dispositif expérimental représentant les différents traitement « lumière » (L+ et L-), « azote » (N+ et N-) et compétition. n correspond au nombre de répétition par traitement (Vernay, 2017).
Des analyses d’azote du sol ont été réalisées en prélevant des échantillons de sol au début et à la fin de l’expérimentation afin d’évaluer la consommation d’azote par la plante. On souhaite également suivre le devenir de l’azote dans la plante. Pour cela il existe une méthode : la méthode isotopique.
Dans la nature, il existe deux isotopes* stables de l’azote : isotope 14N est le plus courant et isotope 15N (Tableau 1).
| Nombre de protons | Nombre de neutrons | |
| Azote 14 | 7 | 7 |
| Azote 15 | 7 | 8 |
L’azote 15 est plus facile à identifier car il est plus lourd que l’azote 14.
Ainsi, on peut mesurer à l’aide d’un spectromètre de masse la quantité d’azote 15 absorbé par chaque organe de la plante.
On pourra donc déterminer où va l’azote une fois absorbé par les racines de la plante.
En juin 2015, les plantes ont été récoltées, lavées, et les différents organes séparés (feuille, tige, racines fines, grosses racines pour le chêne et parties aériennes et parties souterraines pour la canche). Ensuite les échantillons ont été séchés dans une étuve* à 60°C pendant au moins 48h afin d’évaluer la masse sèche* produite (Figure 2).
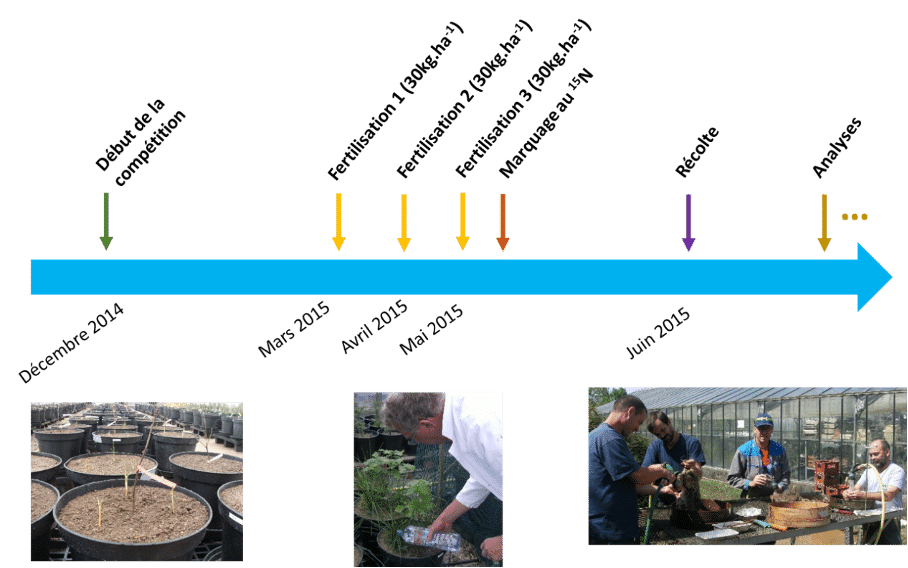
Enfin, les chercheur·se s ont utilisé des indices basés sur les rapports de masses sèches des différents organes afin de quantifier les interactions.
La réponse des plantes dans un milieu correspond en effet aux conséquences des effets de l’environnement et des plantes voisines.
De fait, les chercheur·se s ont calculé, l’intensité des interactions (IINT), soit l’effet absolue de la présence d’une plante voisine sur la réponse d’une espèce cible, et l’importance de ces interactions (IIMP), soit son effet sur la plante cible comparé aux effets dus aux conditions environnementales.
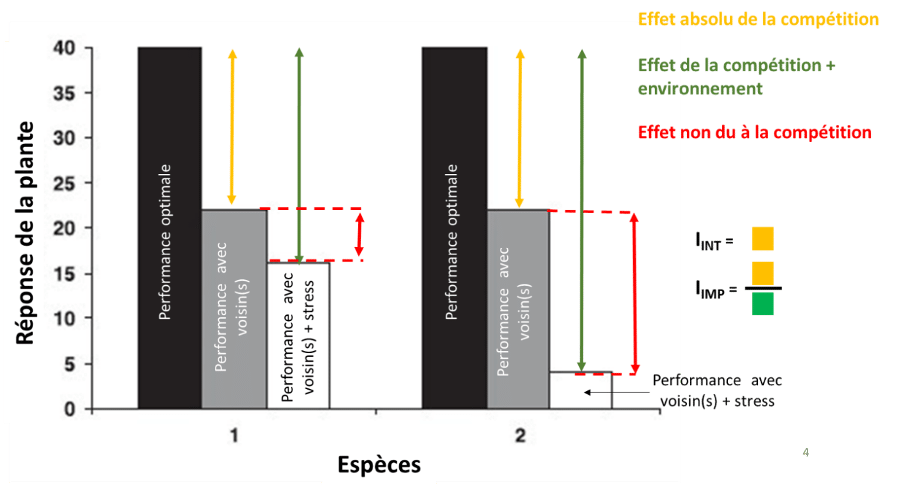
Figure 3 : Explication conceptuelle des indices d’intensité (IINT) et d’importance (IIMP) des interaction modifié de Brooker et al., 2005 et Vernay, 2017)
Si IINT et IIMP sont négatifs, il s’agit d’un effet négatif et donc de compétition d’une espèce sur l’espèce cible, s’ils sont positifs, il s’agit de facilitation.
Plus la valeur absolue* de ces indices est grande plus l’intensité de l’interaction est forte ou pour l’importance, plus l’effet de l’environnement est faible comparé à l’effet de la présence d’un voisin.
III. Résultats
Les résultats montrent d’abord que la biomasse*, notamment aérienne* des chênes sans voisin est plus forte lorsque la lumière et/ou l’azote sont abondants. C’est particulièrement vrai quand les deux sont non limitants. On remarque tout de suite que les plantes sont le moins performantes quand il y a un manque de fertilisant et de lumière et inversement sont le plus performantes quand il y a eu une fertilisation et de la lumière.
Cependant, on remarque que quel que soit l’environnement avec une ressource au moins abondante, le chêne sera moins performant en présence des plants de canches. La lumière joue un rôle important mais la canche avec ou sans chêne montre toujours des performances au moins égales, si ce n’est plus en L+/N+.
Toutefois, dès que les chênes sont en présence de canche, les éléments, azote et lumière, qui lui sont bénéfiques lorsqu’il est seul (sans présence de canche), ne lui apportent aucun avantage. L’interaction azote × diversité, lumière × diversité ont donc un effet négatif fort sur la croissance des chênes (Figure 4). On n’observe pas ces différences dans le compartiment souterrain (données non figurées), les biomasses des racines (fines et grosses) ne sont pas statistiquement différentes malgré les différences de traitement.
A contrario, si l’on se concentre sur les données de biomasse aérienne de la canche, que celle-ci soit en présence de chêne ou non, elle produit la même quantité de biomasse, à une exception près, en présence de grande disponibilité en azote et lumière. Dans cette condition N+/L+, la biomasse aérienne de la canche en présence d’un chêne est même supérieure à celle produite sans ce chêne. De plus, la lumière a un effet positif sur la poussée des racines de canche (données non figurées), la croissance des parties souterraines (racines) est facilitée lorsqu’il y a de la lumière et du fertilisant, elles se développent même encore plus facilement que dans d’autres scénarios.
Ces premiers résultats montrent que le chêne répond principalement à l’interaction des facteurs biotiques et abiotiques lors que la canche répond surtout aux facteurs abiotiques avec peu d’effet de la présence du chêne.
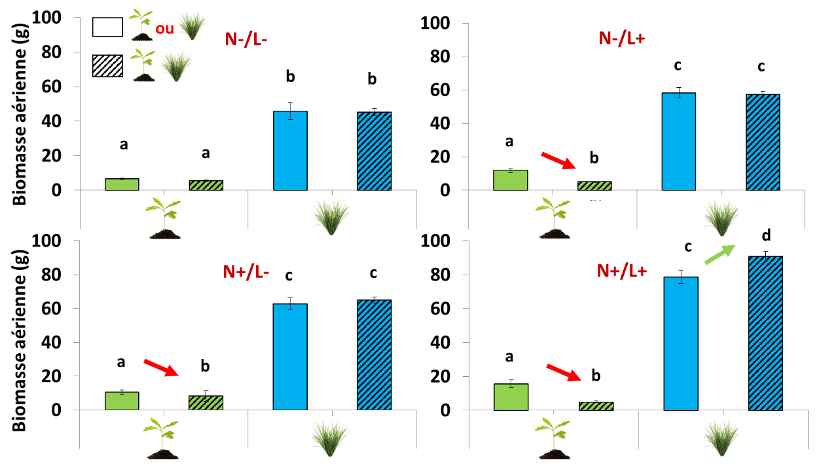
Figure 4 : Production de biomasse aérienne de la canche et du chêne à la fin de l’expérimentation dans les différentes conditions de traitement N × L, en sans compétiteur ou en compétition intraspécifique (barres non hachurées) et en compétition bispécifique (barres hachurées). Les lettres affichées traduisent les différences statistiquement significatives, c’est-à-dire dont le risque que la différence observée soit due au hasard est inférieur à 5%.
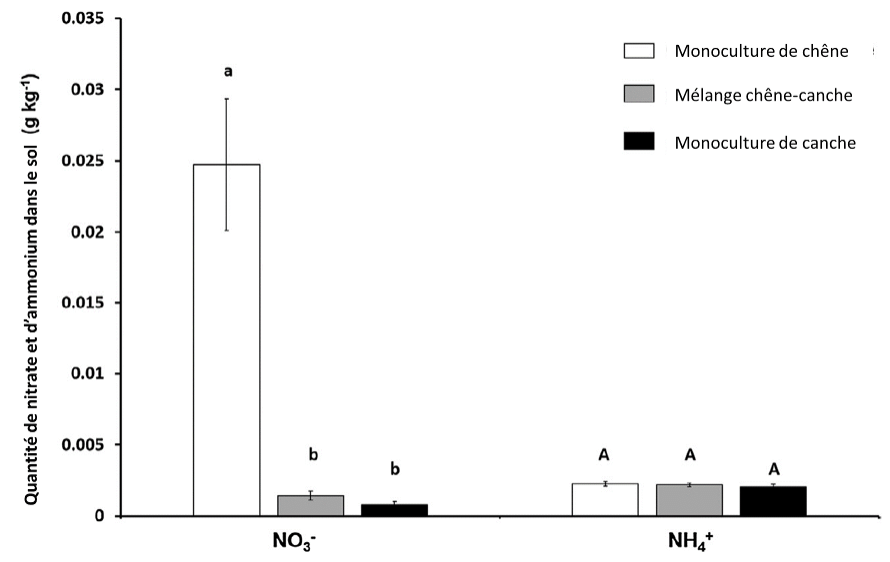
Figure 5 : quantité de nitrate (NO3–) et d’ammonium (NH4+) dans les pots selon leur composition. Les lettres affichées traduisent les différences statistiquement significatives, c’est-à-dire dont le risque que la différence observée soit due au hasard est inférieur à 5%. Les lettres minuscules correspondent à la comparaison liée aux nitrates, indépendamment de la comparaison faite relative à l’ammonium et représentée alors par des lettres majuscules.
Concernant l’utilisation de l’azote, la figure 5 montre la quantité de nitrate et d’ammonium contenue dans le sol selon des conditions de « diversité ».
Les résultats sont donc les suivants :
- La terre des pots de chênes contient 0,025 g/kg de nitrate.
- La terre des pots de mélange chêne et canche contiennent environ 0,002 g/kg de nitrate
- La terre des pots de canche contient environ 0,001 g/kg de nitrate
- La terre des pots de chênes contient beaucoup plus de nitrate que les autres terres des autres pots
- La terre des pots de chêne contient environ 0,0025 g/kg d’ammonium
- La terre des pots de mélange de chêne et de canche contient environ 0,0025 g/kg d’ammonium
- La terre des pots de canche contient environ 0,0025 g/kg d’ammonium
- La terre de tous les pots contient la même quantité d’ammonium
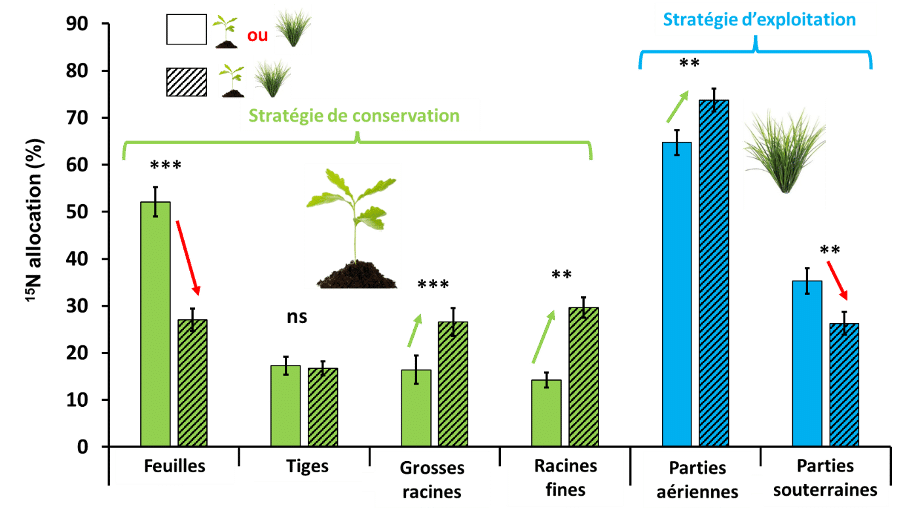
Figure 6 : Allocation du 15N assimilé par la canche et le chêne à la fin de l’expérimentation dans les différentes conditions de traitement N × L, en sans compétiteur ou en compétition intraspécifique (barres non hachurées) et en compétition hétérospécifique (barres hachurées). Les étoiles affichées traduisent les différences statistiquement significatives, c’est-à-dire dont le risque que la différence observée soit due au hasard est inférieur à 5% entre le traitement avec ou sans interaction hétérospécifique. Plus le nombre d’étoile est important moins le risque de se tromper dans l’interprétation de la différence est important. ns signifie non significatif.
La figure 6 parle de l’allocation en pourcentage d’azote dans les organes de la plante. Pour les feuilles, quand le chêne est tout seul, il y a plus d’azote que quand il y a la canche.
La comparaison des tiges de chêne avec ou sans canche n’est pas significative.
Pour les grosses racines, le chêne alloue plus d’azote quand il est en compagnie de la canche. Le résultat est quasiment semblable pour les racines fines.
Pour les parties aériennes, la canche y alloue plus d’azote quand elle est en compagnie du chêne
Pour les parties souterraines, c’est l’inverse, la canche alloue plus d’azote que quand elle est en présence de chêne.
Il y a donc une forte compétition pour l’azote et c’est la canche qui en capture la grande majorité dans le sol. Donc la canche empêche le chêne de croître rapidement.
IV. Discussion
L’un des premiers enseignements de cette expérimentation est que le surplus d’au moins une ressource (lumière ou azote) conduit à la baisse de performance du chêne lorsque celui-ci est en interaction avec la canche. Toutefois, la comparaison entre les performances du chêne à la lumière ou à l’ombre nous montrent un impact fort de la lumière sur la réaction des chênes. Dans le cas où l’azote est aussi présent, l’effet négatif en présence de canche est accentué. L’explication pourrait alors venir d’une photosynthèse plus efficace pour la canche conduisant à une plus forte assimilation* de carbone. La croissance des racines est alors stimulée et également le prélèvement des ressources azotée du sol. Le caractère acquisiteur de la canche se traduit par l’action de capter massivement de la ressource azotée, n’en laissant que très peu de disponible pour le chêne. La canche entre alors dans un « cercle vertueux » car l’azote est un composé essentiel de la Rubisco, une enzyme clé de la photosynthèse ; dans le même temps, le jeune chêne rentre alors dans un cercle vicieux, l’absence d’N pénalisant sa photosynthèse.
La seule condition où la canche n’a pas d’effet négatif sur le chêne correspond au traitement où N et la lumière sont limitants en même temps. Ceci est relativement connu, les espèces très acquisitrices comme la canche sont souvent peu tolérantes aux conditions de stress, se développent moins et laissent donc plus d’espace et de ressources aux espèces plus tolérantes comme le chêne.
Une des découvertes de cet article correspond à la facilitation antagoniste* du chêne sur la canche en abondance de lumière et de N. Nous proposons deux hypothèses pour l’expliquer :
Les racines de chêne peuvent augmenter leur production de rhizodépôt, c’est-à-dire la libération de molécules, tissus dans le sol par leurs racines. Cela peut constituer une source d’azote en plus de celle du sol, rapidement captée par la canche.
Les plantes sont capables de « reconnaitre » les autres espèces en captant les molécules émises dans les rhizodépôts (Semchenko et al., 2014). Si les chênes augmentent leurs rhizodépôts, ce signal pourrait signaler leur présence à la canche et stimuler la croissance de la canche afin de la rendre plus compétitrice face à des individus d’une autre espèce potentiellement utilisatrice des mêmes ressources.
Ces hypothèses explicatives restent toutefois à démontrer dans un contexte semblable.
Cette stratégie permet d’exclure les espèces qui ont besoin de capter du N sans pour autant être efficace dans son assimilation (Tilman, 1982). La canche pourrait alors exclure le chêne, potentiel compétiteur car consommateur de la même ressource en étant plus efficace dans la consommation de N. La réponse du chêne se traduit plutôt dans la redistribution du peu de ressource captée vers des organes de réserves (comme les grosses racines) plutôt que vers des organes de croissance à court terme comme les feuilles. Ceci pourrait lui permettre de mobiliser ces réserves lorsque la pression de compétition sera plus faible (stratégie de conservation).
V. Conclusion et perspectives :
De manière attendue, la canche est extrêmement compétitrice vis-à-vis du jeune chêne, notamment lorsqu’une ressource est non limitante. Plus inattendu, la facilitation antagoniste du chêne sur la canche confirme l’intérêt d’étudier les interactions entre plantes à l’aune de combinaison de facteurs plutôt qu’en se focalisant sur une seule ressource. Les réponses observées seront alors plus proches des conditions naturelles et traduisent la complexité des interactions à considérer, par exemple en contexte de gestion forestière. En effet, nous ne recommandons pas l’utilisation de fertilisant lors de la plantation de chêne en forêt, celle-ci pourrait être contre-productive et favoriser encore plus la canche que les chênes plantés. Il pourrait être plus judicieux de faire constituer des réserves aux chênes en pépinière au préalable (mais cela peut ne pas marcher, Vernay et al., 2018) et d’accompagner cela d’une réduction de la densité de canche, en réduisant la disponibilité en lumière. Cela peut se faire en ne coupant que certains arbres adultes, les autres maintenant un certain niveau d’ombrage.
D’autres ressources comme le phosphate, l’eau pourraient aussi être considérées dans de prochaines études pour compléter cette analyse des interactions entre chêne et canche.
LEXIQUE :
*Assimilation : Transformer, convertir en sa propre substance.
*Biomasse : masse de matière vivante subsistant en équilibre sur une surface donnée du globe terrestre
*Biomasse aérienne : toute biomasse vivante au-dessus du sol, y compris les tiges, les souches, les branches, l’écorce, les graines et le feuillage
*Etuve : Appareil clos et ventilé dont on élève la température afin de sécher les tissus disposés à l’intérieur
*Facilitation antagoniste

*Isotopes : nucléides partageant le même nombre de protons, mais ayant un nombre de neutrons différent.
*Masse sèche : Biomasse des tissus après extraction de l’eau grâce à l’étuve.
*Thalles : pousse aérienne issue de la reproduction végétative des poacées.
*Valeur absolue : La valeur absolue d’un nombre permet de considérer ce nombre sans tenir compte de son signe. Autrement dit, si un nombre x est positif, alors la valeur absolue de x est x, mais si x est négatif, alors la valeur absolue de x est son opposé, soit −x.
Références :
Brooker, R., Kikvidze, Z., Pugnaire, F. I., Callaway, R. M., Choler, P., Lortie, C. J., & Michalet, R. (2005). The importance of importance. Oikos, 109(1), 63‑70.
Semchenko, M., Saar, S., & Lepik, A. (2014). Plant root exudates mediate neighbour recognition and trigger complex behavioural changes. New Phytologist, 204(3), 631‑637. https://doi.org/10.1111/nph.12...
Tilman, D. (1982). Resource competition and community structure. Princeton university press.
Vernay, A. (2017). De la complexité fonctionnelle et écophysiologique des ressources lumière, azote et eau dans le réseau précoce d’interactions entre le jeune chêne (Quercus petraea) et deux Poacées (D. cespitosa et M. caerulea) : Conséquences pour la régénération des chênaies tempérées (AV.tp-00475). Université Clermont Auvergne.
Vernay, A., Malagoli, P., Fernandez, M., Perot, T., Améglio, T., & Balandier, P. (2018). Carry-over benefit of high internal N pool on growth and function of oak seedlings (Quercus petraea) competing with Deschampsia cespitosa. Forest Ecology and Management, 419–420, 130‑138. https://doi.org/10.1016/j.fore...
Ont participé au travail d’écriture de cet article, en collaboration avec Antoine Vernay, chercheur en écologie (par ordre alphabétique) : AIDARA Malick ; BARRIER Lisa ; BEN AYED Maelys ; CHARNOUBI Hafssa ; COLOMBAT Enola ; CORDEIRO-SALIBA Malo ; CROUZOULON Samuel ; CROZET–PONSARD Tony ; DORE Otis ; DUGELAY Camille ; GARCÊS MORGADO Adriana ; GAUVIN Maélia ; GOTTO Keicy ; GRIDEL Swan ; GUENIN ROUBEY Thibault ; LAMBERT Noemie ; LYONNAIS DIOCHON Lisa ; M’MADI Abdourehmane ; MALAGOUEN Mellina ; MALZIEU Djena ; MANE Jordan ; MANGA Merveille ; MEHDI Ahmed ; MOUNIB Meyssane ; MOUSTAÏD Mohamed-Achraf ; OUBRIER Elise ; RIEU Clémence ; SANCHEZ Victoria ; SCHMITT Elea ; SHAK Rayan ; YAHIAOUI Ibrahim ; ZARIOH Marwann ; ZIDANE Lina

Comment citer cet article : Antoine Vernay et la 2nde 2 du lycée Pierre Brossolette (Villeurbanne, FR), Interaction chêne/herbacées : la faim vient en mangeant, Journal DECODER, 2023-06-31
* * *
Vous pouvez retrouver tous nos articles ici !

